如何高效地读完一周的 CNS?
文章来源:未知
作者:老客SEO
人气:13
 2020-02-21 15:32:14
2020-02-21 15:32:14

研究生的主要任务是创造知识,而不是学习知识,因此研究生是没有课本也不需要课本的,取而代之的是对文献的严重依赖。因此,高效地获取文献的要点对于研究生的重要性是不言而喻的。如何锻炼阅读文献的能力呢?如何快速获取一篇文章的要点呢?
我们实验室的答案是 Reference,这是我们实验室长期坚持的一种文献讲解形式,进入实验室的新生需要在每周一的组会上讲解上一周 CNS 杂志上跟生物医学相关的全部文献(如果招了两个人,那么两人轮流讲)。由于 Nature 是北京时间周四凌晨更新,Science 是周五凌晨,而 Cell 是半月刊,因此对于新生而言,需要在三天左右的时间消化 10~25 篇(没有 Cell 的话 10 篇左右,有 Cell 的话再加 10~15 篇)最前沿的学术文章,并且做成 PPT 汇报。有时候新生还有课程和实验,因此时间就显得更紧张,进实验室的第一年,周末基本上都用来看文献了。还好 Reference 并不需要讲的很细,但也得讲成一个自圆其说的小故事。
作为直博三年级的学生,Reference 本来跟我的关系已经不大了,但是为了给新生示范一下如何讲解文献,因此我在本周的组会上汇报了上周 CNS 上的生物医学相关的全部文献,一共 28 篇。我自己看了 3 天(不能算高效,但在高效的路上),感觉看到缺氧,一方面是因为文章虽然都跟生物医学相关,但是涉及的领域实在千差万别,需要一定的基础知识才可以消化;另外还有部分文章真的脑洞大开,思路新奇,几乎超出了我目前的理解范畴,只能读个大概。下面我从切身的体验出发,聊一下自己的一些浅薄的想法。最后,也将我花了很多时间整理的上周的 CNS 生物医学相关的文献要点和 PPT 分享给大家。
做 Reference 的利与弊
(1)锻炼新生阅读文献的能力。正如上面提到了,文献之于研究生,就如枪之于猎人。
(2)让新生尽快掌握基本知识。高中课本上的生物大概念会被细分为一个个领域,这些细分的领域可能是新生此前从来没有思考和接触过的,而大量文献的阅读会迫使新生尽快掌握这些基本的知识,对各个细分领域形成基本的认识。
(3)锻炼演讲的能力。看得明白不等于讲得清楚。首先当然还是要看明白,看明白之后还要思考怎么呈现才能让别人听明白,这又要费一番脑筋。
(4)锻炼做 PPT 的能力。很多人并不重视这一点,我在众多学术会场看到多位大佬的 PPT 实在不堪入目,当然也有一些十分漂亮的。学术 PPT 这个事情,态度完全分立为两派,一派认为学术不需要花里胡哨,内容至上;另一派认为,给人一种舒服的感觉也很重要。学术 PPT 确实需要精简,少一些动画,但是整洁的版面,清晰的图表真的可以增强数据的说服力。
(5)聆听者可快速获取行业最新进展。新生分享的时候,也是导师和高年级学生不断吸收新知识的窗口。
当然,万事都有利弊,做 Reference 也一样。新生一开始可能很难看懂文献,因此会经历一个痛苦的阶段。此外,如果新生准备的 Reference 质量不高(可能是看不懂,也可能是不想看或者没时间看),导师和师兄师姐们也难以获取有用的信息,反而浪费了大家的时间。
如何快速消化一篇文献?
提问是一种非常高效的方法。拿到一篇文献后,不妨先问自己几个问题:(1)这篇文章属于什么领域或方向?(2)解决了什么问题?为什么这个问题这么重要?(3)使用了什么方法和模型?为什么这个方法可以解决这个问题?(4)核心结论是什么?下一步还可以怎么做?
纵向来看,生物过程包括个体层面、组织器官层面、细胞层面、分子层面以及原子层面等。横向切分,这个过程属于什么范畴,比如表观遗传学、免疫、神经生物学、发育生物学等。现阶段的文献,分子层面的机制偏多。由中心法则延展开来,分子无非 DNA、RNA 和蛋白质,这些分子的排列组合起来参与一些特定的过程,比如 DNA 复制、DNA 转录、RNA 翻译、RNA 修饰、组蛋白修饰、非组蛋白修饰等等。当然还有染色体整体层面出发的,比如染色体高级结构调控等。
当然,以上的人为切分其实并不严谨,很多文章涉及了多个过程和多个分子,比如十分火热的早期胚胎发育中表观遗传标志的动态变化,可以认为是发育生物学的典型过程,也可以是表观遗传的经典案例,其中涉及染色质高级结构的动态变化、核小体占位的变化、DNA 甲基化、组蛋白修饰等等,实在太复杂。但是我之所以提出上面的划分,是希望用一些人为的坐标来精确定位这篇文献在生物医学中的具体位置,久而久之,将这些散落的知识点联结起来,最后形成一张知识网络。这大概就是所谓的“connecting the dots”。
按照上面的思路,我对上周的 CNS 文献进行了系统阅读,并整理出了文献的要点。不得不说,有些文章还是没有参透,如有谬误,还请不吝指正。
讲解的文章如下:

大家好,很高兴为大家带来这一期的 Reference。这次一共要讲 28 篇文章,来自不同杂志的文章如果涉及同一领域或者同一话题,我放到一起来讲。所以我先讲 Nature、Science 和 Cell 的封面故事,然后按照文章归类后的顺序讲解。
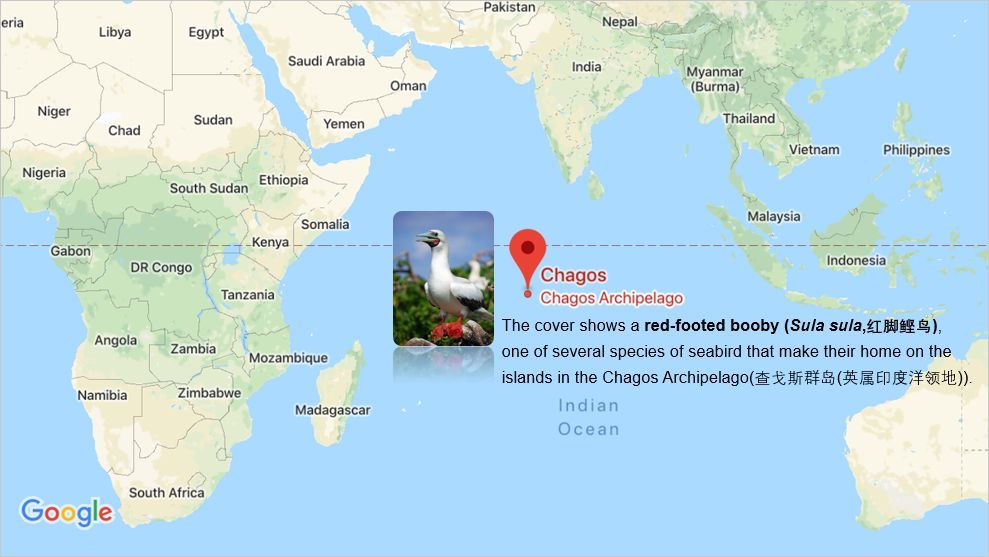
Nature的封面是关于一种印度洋中部的查戈斯群岛上的一种名为“红脚鲣鸟”的海鸟,这种鸟在在矮灌木及小乔木上结群营巢。

Booby /'bu?b?/ 指鲣鸟。鲣鸟不怕人,早期的海员可以轻易地杀死它,因此认为它们笨,故又称之为“笨鸟”,所以 booby 现在也有“呆子、傻瓜”的意思。trap 意为“陷阱、圈套”;这里表示“困境”。这次 Nature 的封面讲的是“鲣鸟的困境”。
鲣鸟的粪便(organicmatter and nutrients)不仅通过循环被海岛的动植物所利用,而且还可以为邻近的珊瑚礁生态系统提供能源,对于珊瑚鱼十分关键。但是随着老鼠的入侵,这种生态平衡被破坏。老鼠的出现后,鲣鸟蛋面临很大威胁,鲣鸟的数量下降,营养物质也随之显著下降。因此,消灭海岛上的老鼠可以纠正这种被破坏的平衡,最终帮助珊瑚礁的恢复。

Science封面提到了两个核心概念:中微子和耀变体。
所谓“中微子”,“中”是指电中性,“微”是指个头小,而且质量非常轻(有的小于电子的百万分之一)。中微子以接近光速运动,可自由穿过地球,每秒钟有 6500 万个中微子穿过我们的指甲盖。中微子与其他物质的相互作用十分微弱,号称宇宙间的“隐身人”,可能是现在唯一一种已探测到的暗物质。中微子来源包括太阳中氢原子的核聚变,宇宙射线与地球空气粒子的碰撞以及核裂变。
耀变体是一种密度极高的高变能量源,被认为是一种超大质量黑洞。黑洞的融合可以释放出中微子。

为了探测中微子,科学家们在南极洲建立了“冰立方中微子观测站”。一些直径约为 35cm 的,圆形的数字光模块被放置到冰面以下 2.5 千米的地方。超过 5000 个数字光模块形成了一个巨大的一平方公里的探测器。这些数字光模块可以检测到高能中微子与冰相互作用中释放的极其微弱的光。最终科学家们利用“冰立方”检测到了来自耀变体的中微子。

Cell 封面讲的是一种轮藻,它被认为是陆地植物的进化史的祖先。科学家们绘制了布氏轮藻的基因组草图,揭示了一些对植物陆地化十分关键的进化特征。封面显示的是布氏轮藻的显微图,图中包含两个椭圆形的卵原细胞(雌配子囊),以及两个相关的小的,圆形的雄体(雄配子囊)。芦苇芽植物门是陆生植物的共同祖先,包括苔藓、蕨类植物、裸子植物和被子植物,而基因组的数据显示轮藻与芦苇芽植物门的生物学基础很像。

这是 Nature本期的全部文章,要讲的文章已经标注,其中蓝色的是跟生物相关的,棕色的是一篇天文学的,稍作科普。


这是 Cell 本期的全部文章,封面的文章刚才已经介绍之前,另一篇棕色标注的不讲,其余的全部讲解。

第一篇文章来自 Nature,讲的是一个星际外天体的非引力加速。这个星际外天体的名字非常独特,叫 Oumuamua,国内官方的翻译为“奥陌陌”,夏威夷语中意为“远方的信使”。1I 中字母 I 表示“星际的(Interstellar)”,1 表示第一个;这个星际外的天体长约 400 米,宽约 40 米,呈现雪茄形,如图中所示的样子。2017 年 10 月 19 日被发现,这是人类首次在太阳系内发现系外天体。

科学家们对奥陌陌的轨迹进行了模拟,发现它从太阳系之外进入太阳系,随后又快速远离太阳系。因此人们猜测,这是否是太阳系外文明发射的一种探测器?
生物医学相关的论文解读如下:
主题一:相分离
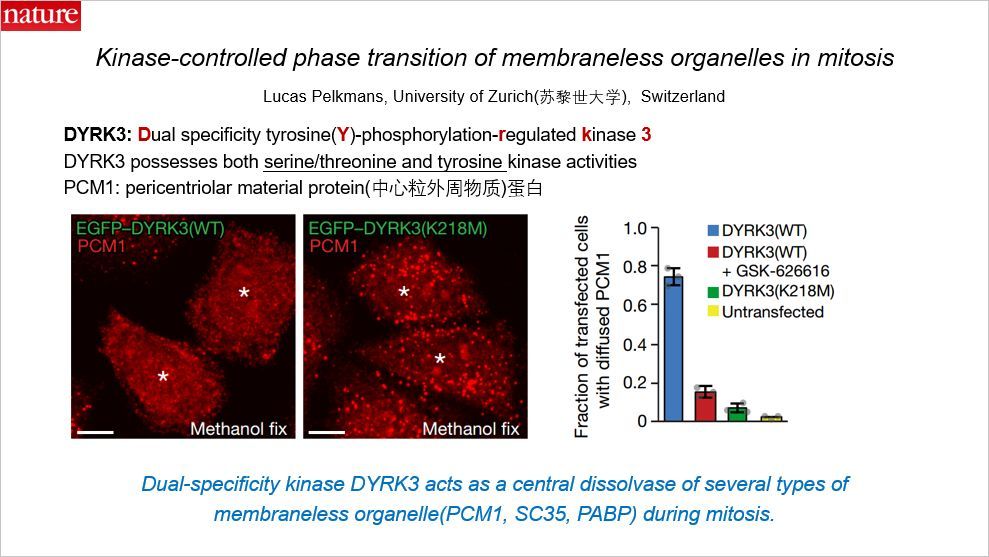
这篇 Nature 涉及的主题是近几年十分火热的“相分离”领域,作者重点研究的是有丝分裂过程中由激酶控制的相分离过程,本文通讯作者来自瑞士苏黎世大学。
文章聚焦的蛋白是 DYRK3: Dual specificity tyrosine-phosphorylation-regulated kinase 3。DYRK3 的双重特异性是指其同时具有丝氨酸/苏氨酸和酪氨酸激酶活性。而可以形成相分离的蛋白包括中心粒外周蛋白 PCM1
当 DYRK3 功能缺失型突变后,PCM1 更容易形成相分离。注意右边的柱状图的纵轴显示的是溶解的蛋白。相分离后,溶解的蛋白减少。类似地,用 DYRK3 的抑制剂 GSK-626616 可以得到类似的结果。在有丝分裂过程中,激酶 DYRK3 作为一些无膜细胞器的核心的溶解酶。这些无膜的细胞器除了 PCM1 之外,还包括剪接颗粒(splicing-speckle)的marker SC35 以及应激颗粒(stress-granule)的 marker PABP。
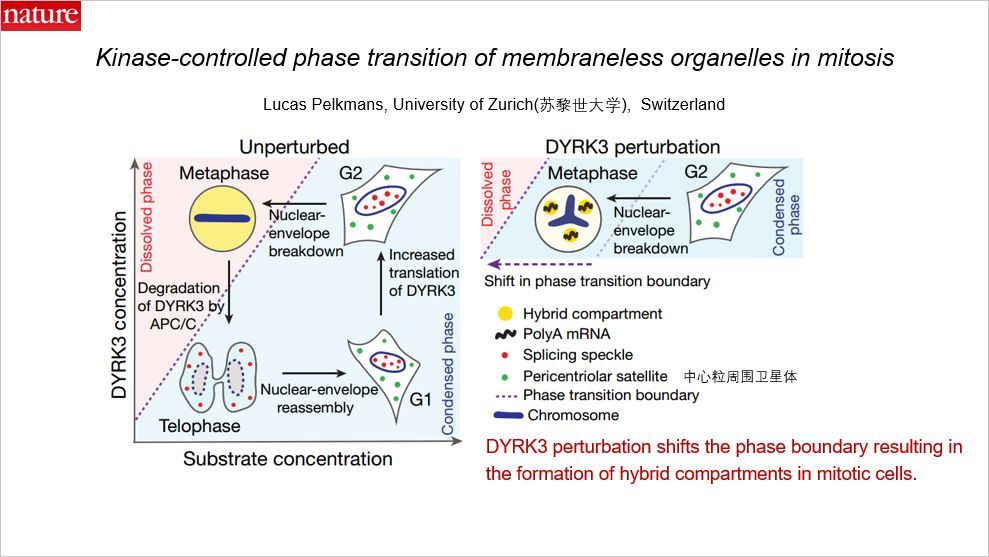
放眼细胞中:在正常细胞周期下,在 G1 期细胞中存在一些液相的无膜细胞器,比如剪接颗粒(splicing-speckle)和中心粒周围卫星等。在 S 期 DNA 复制, DYRK3 增多,一旦进入分裂期,这些颗粒溶解,形成一种混合的液相。而在有丝分裂后期,DYRK3 被降解,在子细胞中又可以形成相分离。而一旦 DYRK3 被扰乱,即使进入细胞分裂期,细胞中还是会存在着相分离现象。因此,DYRK3 的扰乱导致了相分离边界的移动,导致在有丝分裂细胞中混合区室(hybrid compartments)的形成。
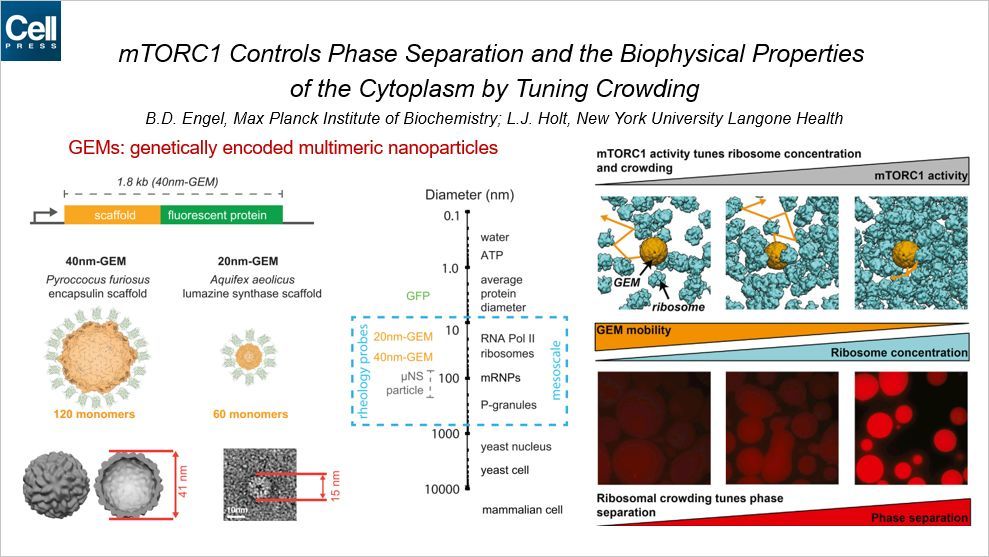
另一篇是相分离的文章来自德国马普所和纽约大学朗格尼医学中心。德国马普所是研究相分离的重镇,首次提出 P-颗粒的相分离概念的就是马普所的研究员。
细胞内大分子的拥挤程度对于多种生物过程十分关键,但是其背后的调控机制还不是很清楚。为了研究这个问题,作者首先构建了一种遗传学编码的多聚物纳米粒子,简称 GEMs。GEM 可以被定制为不同的尺寸和形状,用来检测细胞内大分子的拥挤程度。比如 40nm 的 GEM 可以用来检测 P-颗粒、mRNPs、核糖体等大分子的密度。
通过研究发现,mTORC1 的活性可以调控核糖体的密度和拥挤程度。当核糖体越密时,GEM 运动就越困难。而核糖体的拥挤程度又可以调控相分离,核糖体密度越高,相分离现象越明显。所以,总的来说,mTORC1 可以通过控制核糖体的密度而控制相分离。
主题二:Genomics and its related events
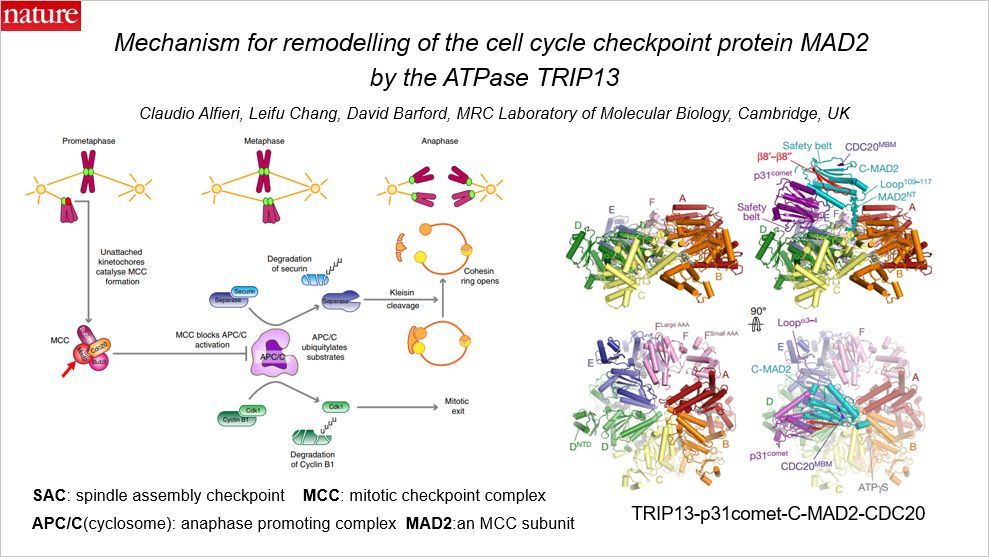
这篇 Nature 涉及有丝分裂检查点,在有丝分裂中期到后期之间存在一个检查点。这篇文章揭示了 ATP 酶 TRIP13 介导的细胞周期检查点蛋白 MAD2 的变构。本文作者只有三名,均来自剑桥 MRC 实验室。
有丝分裂过程中基因组稳定性的维持是纺锤体组装检查点(SAC,spindle assembly checkpoint)通过它的效应蛋白有丝分裂检查点复合物 MCC 调控的。MCC 本身可以抑制后期促进复制物APC/C 复合物。MAD2 是 MCC 复合物的一个亚基。未经检查的着丝粒通过催化 MAD2 β-折叠的拓扑结构变化而控制 MCC 组装,因此产生激活的关闭状态的MAD2(C-MAD2)构象。因此不能进入细胞分裂后期。MCC 的解聚对于 SAC 失活和染色体分离是必须的。MCC 的解聚是一种 ATP 依赖的过程,由 AAA+ATPase TRIP13 驱动。 TRIP13 与一种 SAC 拮抗剂 p31comet 组合起来,可以将 C-MAD2 重构为失活的打开的状态(O-MAD2)。
本文解释了 TRIP13-p31comet 解聚 MCC 的机制。TRIP13-p31comet-C-MAD2-CDC20 复合物的冷冻电镜结构揭示了 p31comet 招募 C-MAD2 到 TRIP13 六边形环中特定的位点,将 C-MAD2 的 N 端(MAD2NT)插入到 TRIP13 的轴向孔中,扭曲 TRIP13 环以启动结构的重塑。
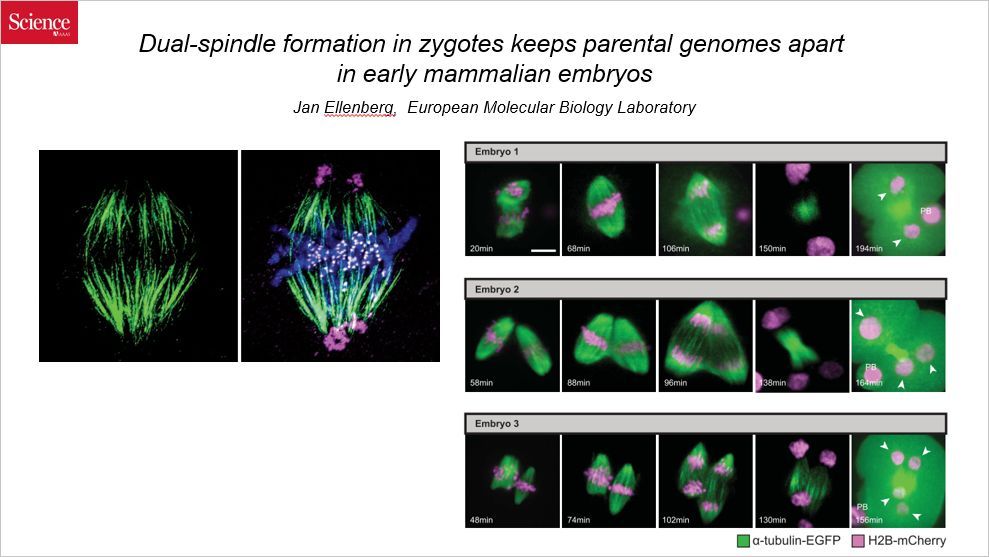
这篇 Science 是关于是关于哺乳动物早期胚胎卵裂过程中两个纺锤体形成的。通讯作者来自欧洲分子生物学实验室。
此前人们认为,精子和卵子在结合以后,来自父亲和半套染色体和来自母亲的染色体组装成整套染色体,在卵裂过程中作为整体复制和分离;而这篇文章发现,来自父亲的染色体和来自母亲的染色体“貌合神离”,在染色体分离的时候“各自为政”,使用两套纺锤体分离,而且两套纺锤体还不同步,因此有可能导致多核的产生。
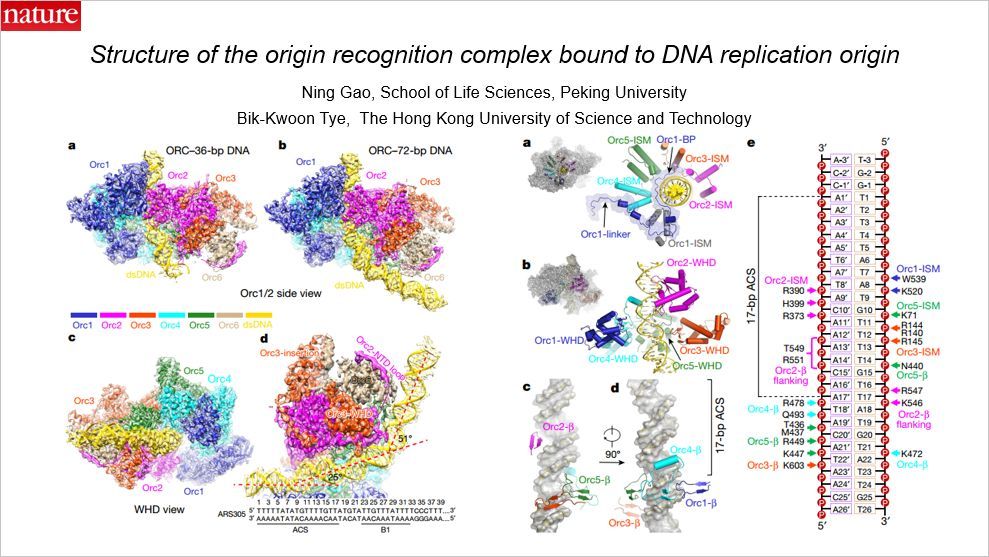
这篇 Nature 解析了 DNA 复制起始复合物的结构,文章通讯作者为北京大学生科院的高宁以及香港科技大学的 Bik-Kwoon Tye。
DNA 起始复合物拥有六个亚单元,这篇文章用冷冻电镜解析了酿酒酵母 ORC 结合到一个 72bp 起始 DNA 序列的冷冻电镜结构,分辨率达到的 3Å。
这篇文章是关于 DNA 复制的,通讯作者来自丹麦。研究发现高速的复制叉进展诱导 DNA 复制应激和基因组不稳定性。准确的 DNA 复制需要严格的调控机制以保证基因组的完整性。细胞的 S 期,数千个复制起始位点(origins of replication)有序地被激活,复制叉的速度需要合适,一方面需要完整地复制 DNA,另一方面还需要跟细胞周期同步。当细胞遇到应激时,复制压力可以导致复制叉的暂停,产生基因组的不稳定性,但其背后的原因还不清楚。
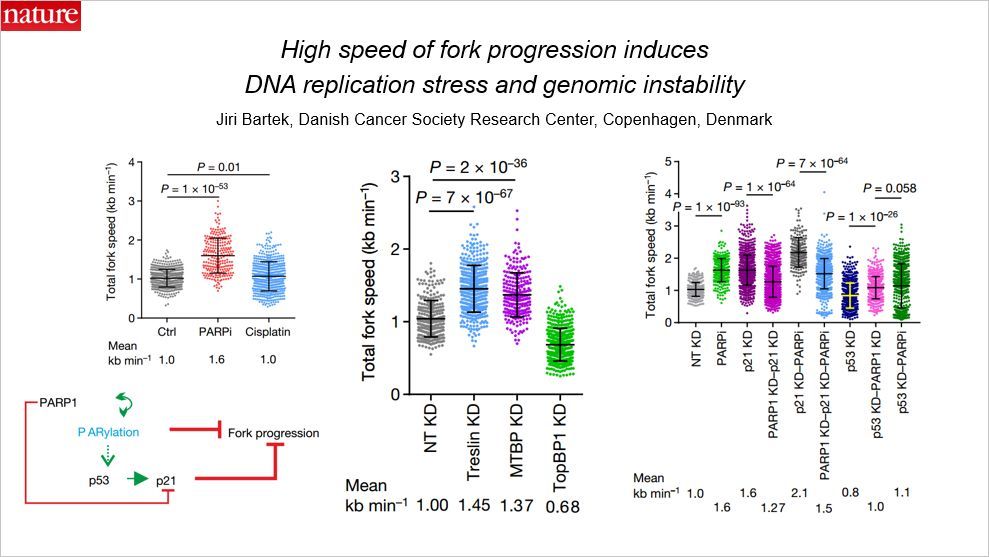
本文显示,PARP(聚(ADP-核糖)聚合酶)的抑制加快了复制叉延伸的速度,但并不会导致复制叉的暂停,这与此前广泛接受的模型正好相反的。此前人们认为,PARP 的抑制剂会诱导复制叉的暂停和崩溃。复制叉的异常加速了大约60%,会导致 DNA 损伤。
treslin 或者 MTBP 蛋白(这些蛋白参与复制的起始)的敲除也可以提升复制叉的速度,超过了可接受的阈值,诱导 DNA 损伤响应通路。在机制上,多聚(ADP-核糖基化)以及 PCNA 的相互作用蛋白 p21Cip1 对于复制叉的推进十分关键,它们可作为复制叉速度的抑制因子,但受 PARP1 和 p53 蛋白控制。在复制叉水平,多聚(ADP-核糖基化)可以作为复制压力的一个感应因子。在 PARP 抑制过程中,诱导复制叉阻滞以及被正常解决或修复的 DNA 损伤维持不被复制机器识别的状态。
这项发现有助于更好地理解复制叉的速度控制机制,在基因组稳定性或者肿瘤治疗中具有潜在的应用价值。
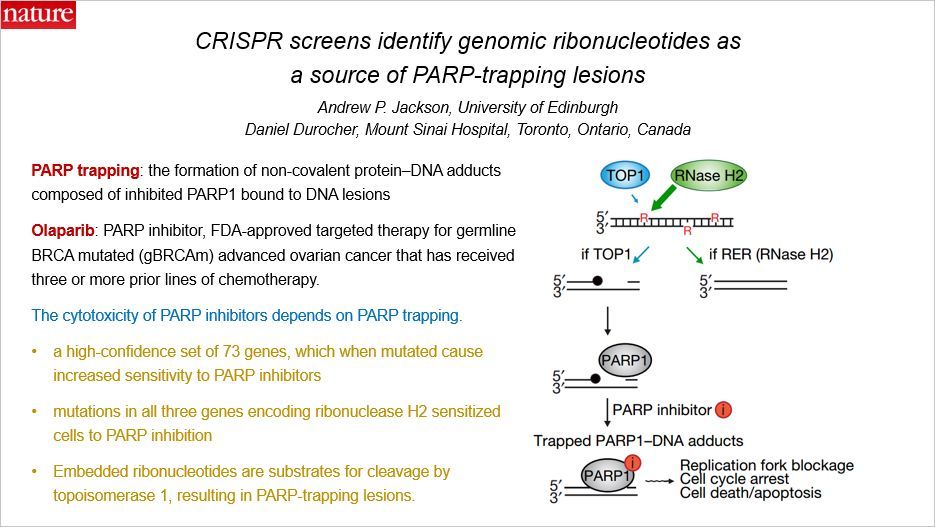
多聚(ADP-核糖)聚合酶(PARP)是 DNA 损伤的感应器,对于 DNA 损伤修复和基因组稳定十分关键。PRAP 家族蛋白包含 18 个成员,其中 PAR1 和 PARP2 研究的最多。近些年,PARP 抑制剂也被广泛应用来对抗肿瘤,比如奥拉帕尼,这是一种临床上获批的 PARP 抑制剂。
PARP 陷阱(PARP-trapping)是由抑制后的 PARP1 结合到 DNA 损伤位点形成的非共价的蛋白质-DNA 加合物。BRCA1 和 BRCA2 缺陷的细胞对 PARP 抑制剂很敏感,而PARP抑制剂的细胞毒性依赖于PARP陷阱。为了鉴定这种损伤的特性和PARP陷阱的细胞结果,作者利用 CRISPR 筛选来鉴定介导细胞抵抗奥拉帕尼的基因和通路。
文章发现了 73 个高可信度的基因,当它们突变后,细胞对 PARP 抑制剂的敏感度升高。编码核糖核酸酶 H2 的三个基因的突变让细胞对 PARP 抑制敏感。存在编码核糖核酸酶 H2 缺陷的,对 PARP 抑制剂低敏感的细胞,其核糖核酸切除修复受损。嵌入的核糖核苷酸,在核糖核酸切除修复存在缺陷的细胞中很常见,是拓扑异构酶 1 进行剪切的底物,进而导致了 PARP-陷阱损伤,影响 DNA 复制和危害基因组的完整性。基因组的核糖核苷酸是目前还没有引起重视的 PARP-陷阱 DNA 损伤的来源,而这些发现可能应用到治疗 RNASEH2B 频繁缺失的转移性的前列腺癌和慢性淋巴白血病中。
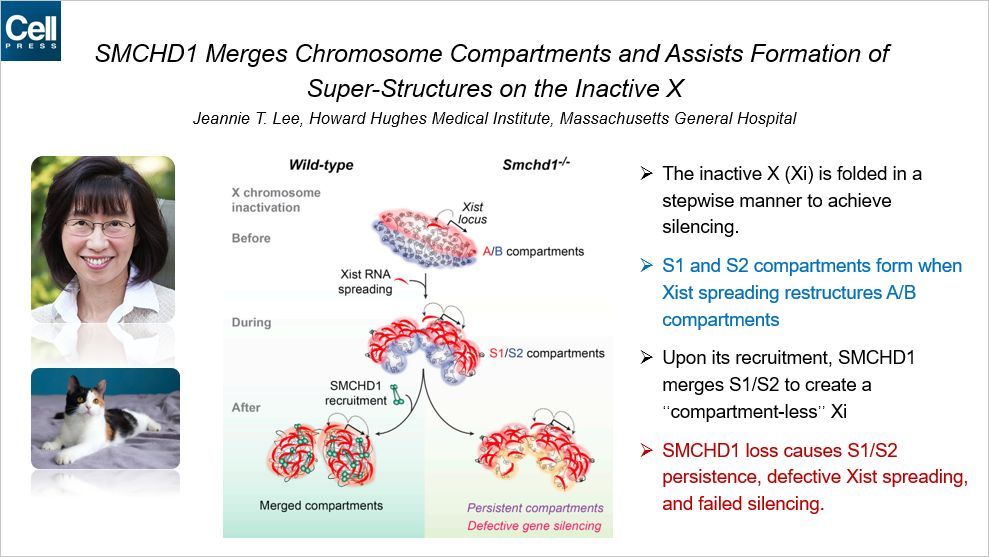
这篇 Cell 是关于 X 染色体失活的,SMCHD1 可以融合染色质区室,有助于在失活的 X 染色体上形成超级结构。本文通讯作者是 HHMI 和哈佛的 Jeannie Lee,Jeannie Lee 是研究 X 染色体失活的专家,她最早发现了 X 染色体失活中心(Xinactivation center, XIC),并用 lncRNA Xist 包裹失活的 X 染色体来解释 X 染色体失活现象。生活中关于 X 染色体失活的例子很多,最著名的当属三色猫了。因为控制毛色的基因位于 X 染色体上,在胚胎发育的早期,细胞中 X 染色体随机失活,因此有的细胞中带有黄毛基因,有的携带棕色基因,因此猫身上呈现出不同的色块。
回到本文,Jeannie Lee 发现,在正常生理条件下,失活的 X 染色体逐步折叠以实现基因沉默。而随着 XistRNA 的传播,它可以将正常的染色质高级结构 compartment A/B 重新构造为 compartment S1/S2。随后,SMCHD1 可以将这两个 compartment合并,形成一个“少compartment”的失活的X染色体。而当 SMCHD1 双敲以后,会导致 compartmentS1/S2 不能合并,而且 Xist 无法传播,最终导致这条 X 染色体不能正常失活。
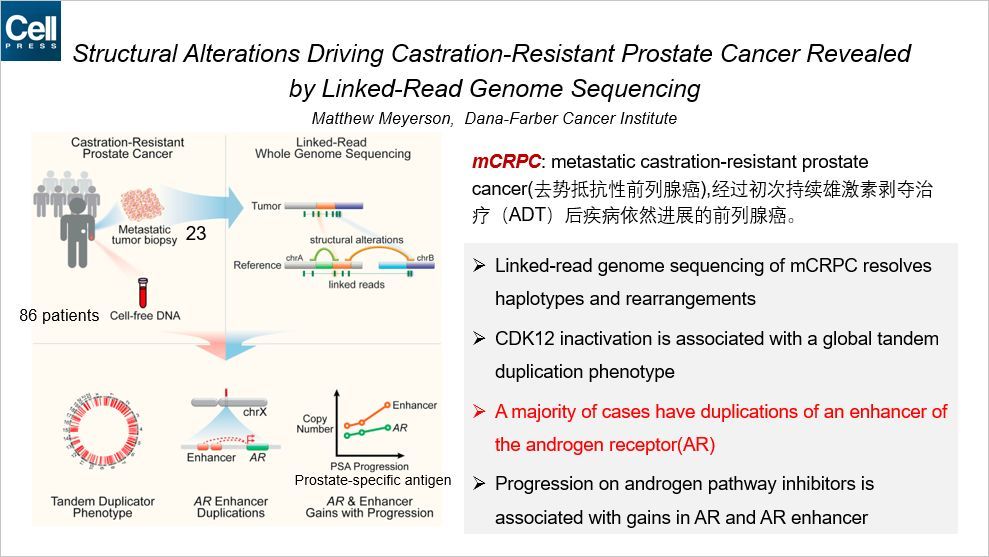
这篇文章是关于染色质重排介导的前列腺癌的,通讯作者来自 Dana-Farber 癌症研究中心。作者同时检测 23 个转移的肿瘤活检样本和来自 86 个病人的 cfDNA。通过 Linked-read 基因组测序解析了 mCRPC 的染色质重排事件。CDK12 的失活与全基因组的串联重复的表型相关。在大部分样本中发现了雄激素受体上游一个增强子的重复。随着前列腺特异性抗原(PSA)的进展,AR 和它增强子的扩增也增多。
主题三:RNA biology
讲完了基因组及其相关的事件,我们转换到 RNA。
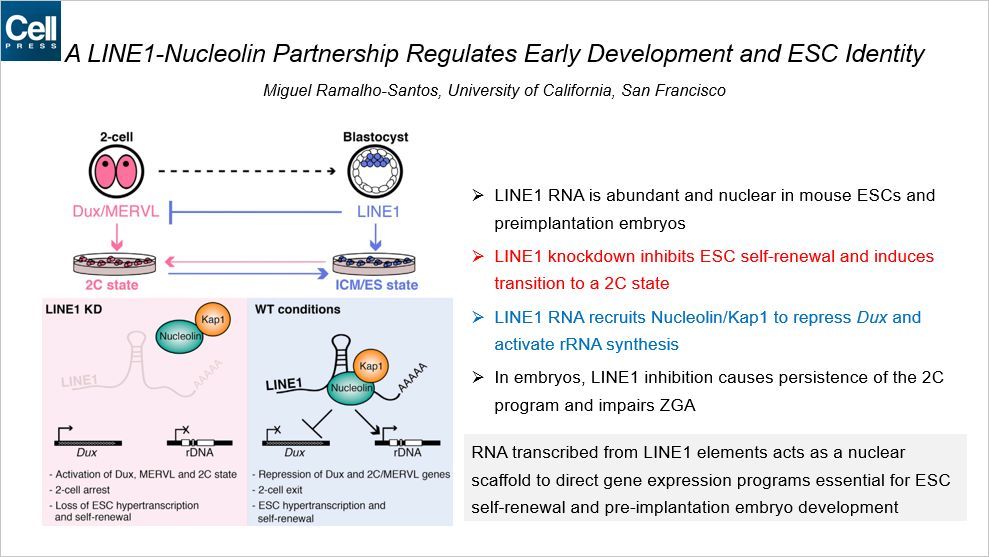
这篇 Cell 是关于 LINE1 RNA 的,通讯作者来自 UCSF。
LINE1 是哺乳动物体内分布广泛的重复序列元件,其功能还不是很透彻。本文发现 LINE1 RNA 在小鼠胚胎干细胞和着床前的胚胎的细胞核中有很多。LINE1 敲低抑制了 ESC 自我更新,诱导向 2-细胞状态的状态。LINE1 RNA 招募核仁蛋白(Nucleolin)/Kap1来抑制 Dux 和激活 rRNA 的合成。在胚胎中,LINE1 抑制导致 2-细胞状态的维持,影响了合子基因组激活。
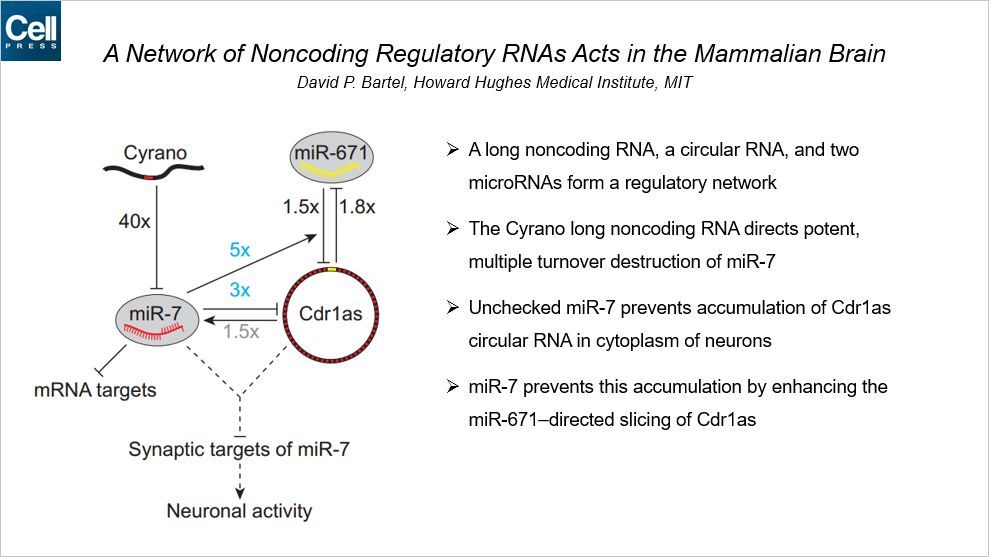
细胞里面存在多种多样的非编码 RNA,比如 miRNA、lncRNA、circRNA 等。这篇 Cell 发现了不同类型的非编码 RNA 之间存在 crosstalk,而且可以形成一个调控网络。通讯作者来自 HHMI 和 MIT。
这个调控网络包含了一个 lncRNA,一个 circRNA 和两个 miRNA。首先 lncRNA Cyrano 可以抑制 miR-7。而不受抑制的 miR-7 可以防止 circRNA Cdr1as 在神经元胞浆中的聚集。miR-7 又可以通过增强 miR-671 介导的 Cdr1as 的沉默作用来防止它的聚集。

这篇 Cell 来自于加州理工学院的华人科学家蔡龙教授。他 2001 年毕业于哈佛大学,2006 年在哈佛获得博士学位,博士期间在哈佛谢晓亮的实验室。
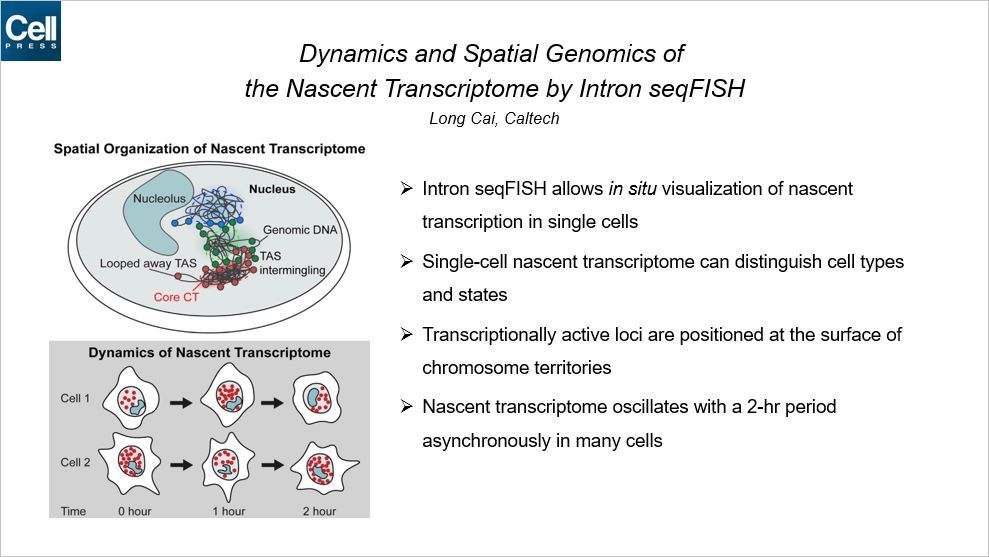
这篇文章建立了一个多路复用的单分子原位方法,通过内含子 seqFISH 结合mRNA、lncRNA seqFISH 和免疫荧光,在单个细胞中允许 10421 个基因在它们新生的转录活性位点成像。在小鼠胚胎干细胞和成纤维细胞中,这种转录组谱系方法可以鉴定不同的细胞类型和状态。RNA 合成的新生位点倾向于定位到染色质域(chromosome territories)的表面,在单个细胞中,它们的组织形式是高度变化的。在单个小鼠的胚胎干细胞中,2h 时间内整体的新生转录本异步震荡,这种情况也适用于成纤维细胞。
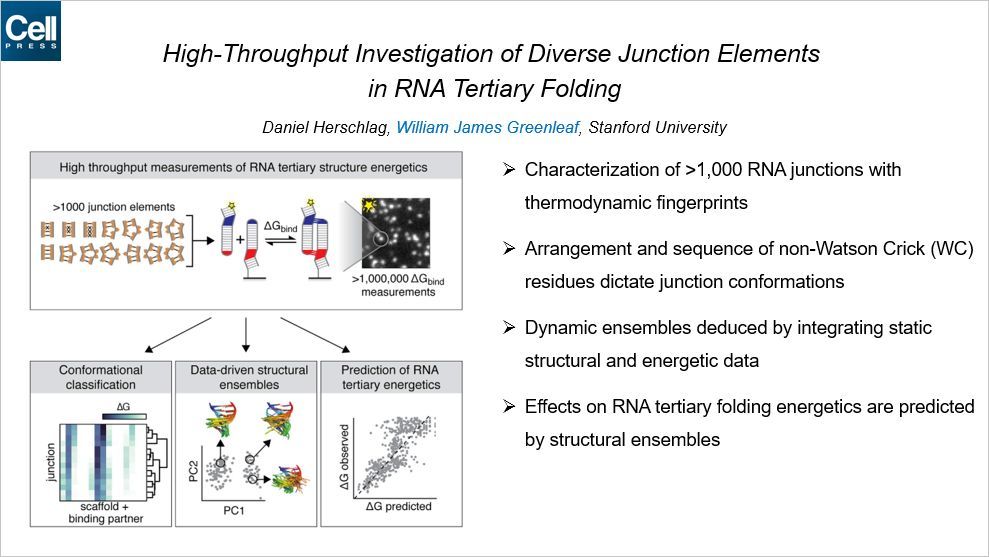
这篇 Cell 来自斯坦福大学的 William Greenleaf,我们对 William Greenleaf 已经不再陌生了,他是 ATAC-seq 的创建者,而且去年 9 月底来参加我们的国际表观会议,我当时还去接待的他。
在这篇文章中,Greenleaf 开发了一种用来预测 RNA 三级结构的模型。首先,通过一种叫做热动力指纹(thermodynamic fingerprints)的方法,他们鉴定了超过 1000 个 RNA 连接点。RNA 与 RNA 之间通过双向连接,一旦连接后,可以释放荧光。非沃森-克里克序列的重排和序列指导了联结的构象。通过整合静电结构和能量数据推演出 RNA 的动态组装。RNA 三级结构折叠的能量的影响可通过结构组装而预测。
主题四:Immunology
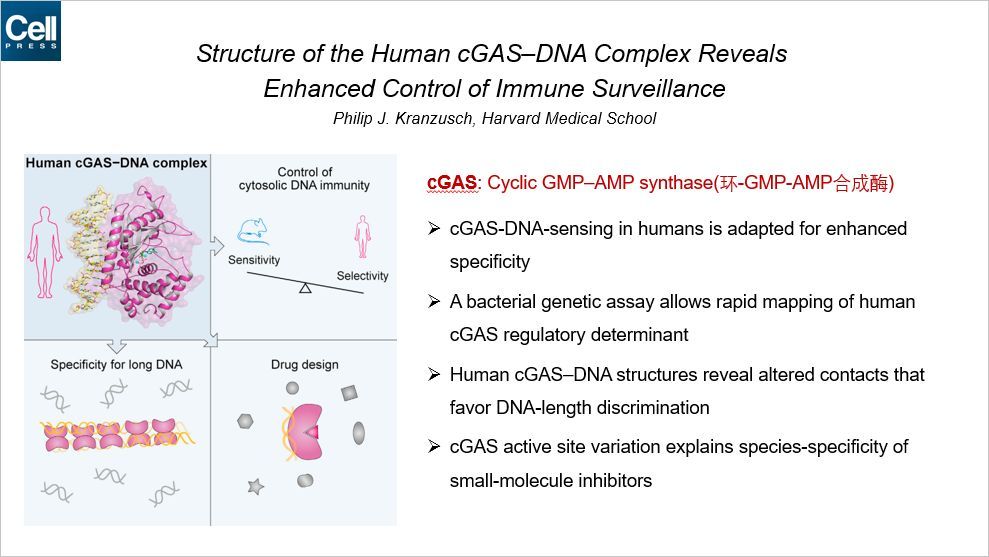
这篇文章通过 cGAS-DNA 复合物的结构揭示了免疫监控的增强控制。通讯作者来自哈佛医学院。cGAS 识别胞浆中的 DNA 是细胞的一种重要的免疫响应途径,用来监控病原体的复制、细胞压力和癌症等。人类相比于其他的哺乳动物,cGAS 的环-GMP-AMP 的酶活显著底下的。
本文发现了人类 cGAS 对 DNA 长度具有更强的特异性,这限制了其酶活。通过细菌实验,将人类 cGAS 调控的决定因素精确到两个氨基酸上。人类特异性的氨基酸替换对于感知长的 DNA(大于45nt,而老鼠可以感知 20nt 的)是必要而且充分的。晶体结构揭示了改变了的相互作用更倾向于 DNA 长度的选择。这些结果揭示了在人类中感知特定长度 DNA 的机制,使得基于 cGAS 疗法成为可能。
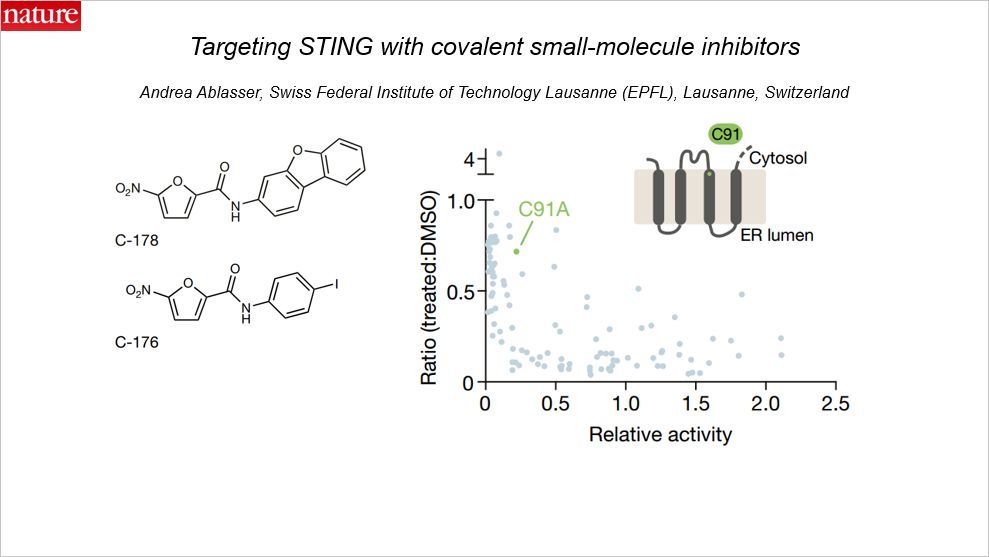
天然免疫通路的异常激活与一系列疾病相关。天然免疫通路的分子机制的进展有助于研发相应的靶向治疗策略,但是要靶向天然免疫通路中的特定蛋白还很困难。这篇 Nature 报道了高效能的,高选择性的 STING 蛋白的小分子拮抗剂。STING 是细胞外的 DNA 响应通路的核心分子。
在机制上,这种被鉴定出来的化合物可以共价靶向预测的跨膜的第 91 位的半胱氨酸残基 ,因此可以阻断激活诱导的 STING 的十六烷酰化。利用这些抑制剂,发现 STING 的十六烷酰化对于它组装进高尔基体中多聚复合物十分关键,反过来,又可以招募下游的信号分子。在人和小鼠细胞中,这种被鉴定的小分子以及它们的衍生物降低了 STING 介导的免疫炎性因子的产生。另外,这些小分子拮抗剂降低了小鼠中自身免疫性疾病的病理特征。总的来说,这项工作揭示了 STING 被药物抑制的机制,展现了通过靶向 STING 而治疗自身免疫性疾病的潜力。
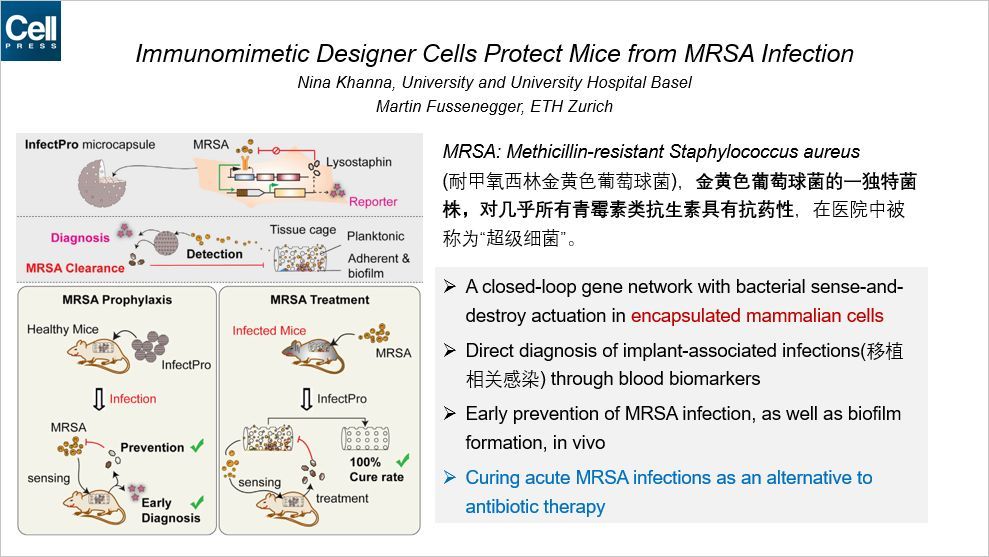
这篇 Cell 构建了一种基因工程化的细胞,可以对抗超级细菌的感染,本文通讯作者来自瑞士。这种人为构造的细胞具有闭合的基因环路,具有潜在的临床应用价值。
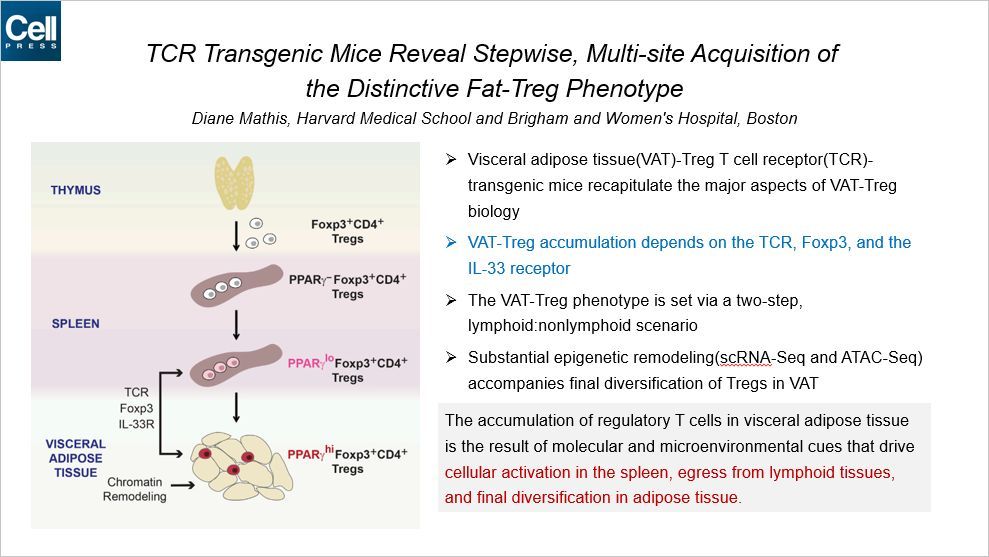
这篇 Cell 利用转基因小鼠揭示了内脏脂肪组织中调节性 T 细胞多样性的机制。众所周知,淋巴细胞不仅仅存在于淋巴组织中,而非免疫组织中也存在多种多样的淋巴细胞,比如肝脏、肾脏等,这类细胞称为“组织驻留的淋巴细胞(Tissue-Resident Lymphocytes)”。本文聚焦的是内脏脂肪组织(VAT)中的辅助性 T 细胞(Treg)多样性形成的机制。文章发现了一些对 VAT-Treg 聚集十分关键的因子,比如 TCR 和 Foxp3 和 IL-33 等。
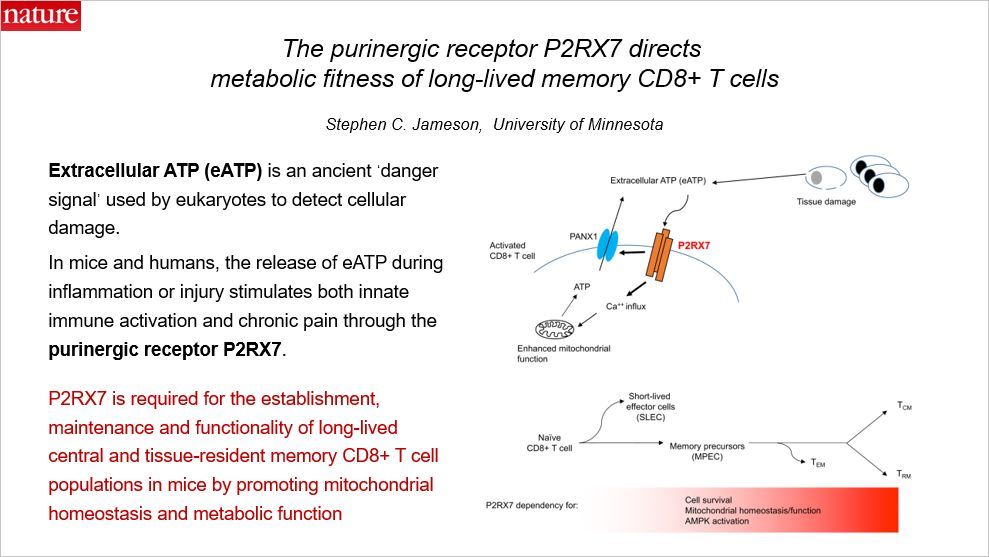
这篇 Nature 揭示了嘌呤受体 P2PX7 指导长期存在的 CD8+ T 细胞的代谢。关于 T 细胞的分化和来源,参见此前的科普文“记忆 T 细胞的前世今生”。这篇文章发现嘌呤受体 P2RX7 对于长期存在的和组织驻留的 CD8+ T 细胞的产生、维持和发挥功能都十分重要,而且是通过线粒体稳态介导的。
主题五:Signaling pathway
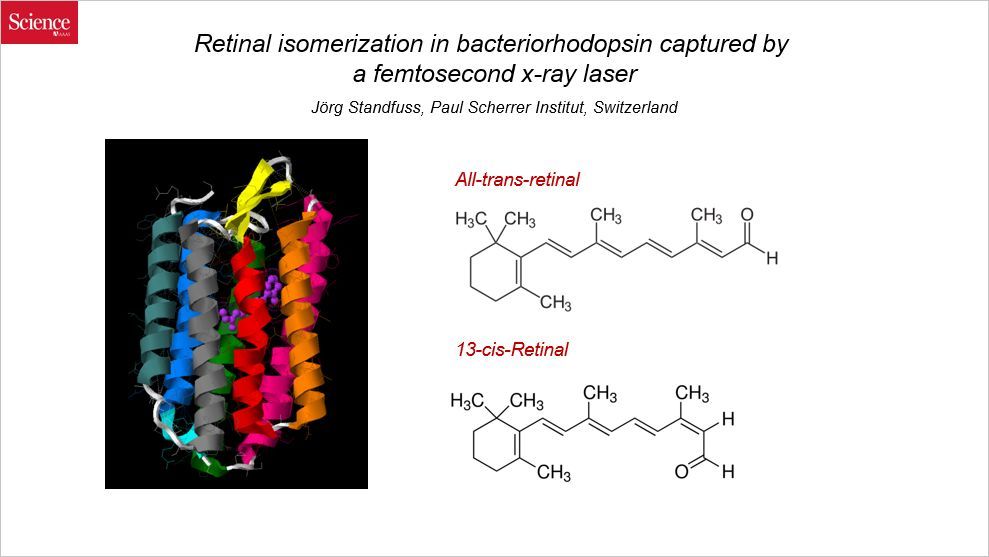
很多生物都有光敏感性的蛋白,比如植物利用叶绿素将光能转化为化学能,而动物则利用视紫红质。人类的视紫红质分布在视杆细胞中,可以将光能转变为电信号,是我们视觉形成的关键环节;而细菌不需要视觉,细菌的视紫红质作为质子泵,在光能的刺激下传导质子。本文讨论的是细菌视紫红质(bacteriorhodopsin),通讯作者来自瑞士保罗谢勒研究所。
在结构上,视紫红质由视黄醛和视蛋白结合而成。视蛋白是一种 G-蛋白偶联受体,它包含 7 个跨膜结构域;而视黄醛是一种小分子,包裹在视蛋白里面。光可以改变视黄醛的构象,进而刺激质子的跨膜运输。因此研究视黄醛的超快的异构化对于理解视黄醛介导的光化学反应具有重要意义。
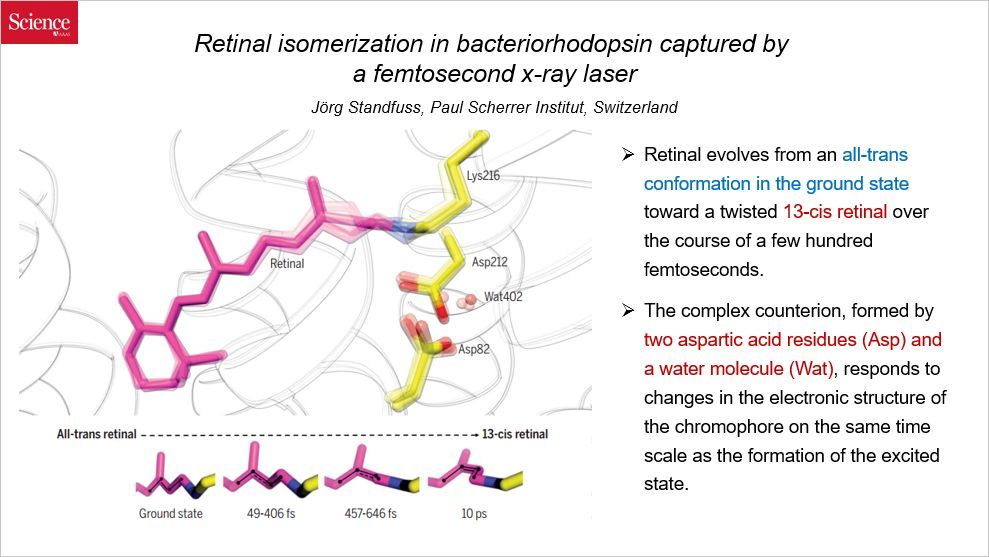
本文利用 X 射线,在亚皮秒(皮秒是1×10-12s,比皮秒还小)尺度,研究了细菌视紫红质中视黄醛的结构动态。一系列将近原子分辨率的结构快照揭示了蛋白结合口袋中全反式视黄醛构象状态如何在数百飞秒(1×10-15s)内通过一个几何扭曲转变为 13-顺式结构。由两个天冬氨酸残基和一个水分子形成的平衡离子,也可以在同样的时间尺度响应视黄醛的结构变化,这说明天冬氨酸残基和水分子可作为超快集体动态作为细菌视紫红质这种立体选择性和高效光化学反应的关键小平面。
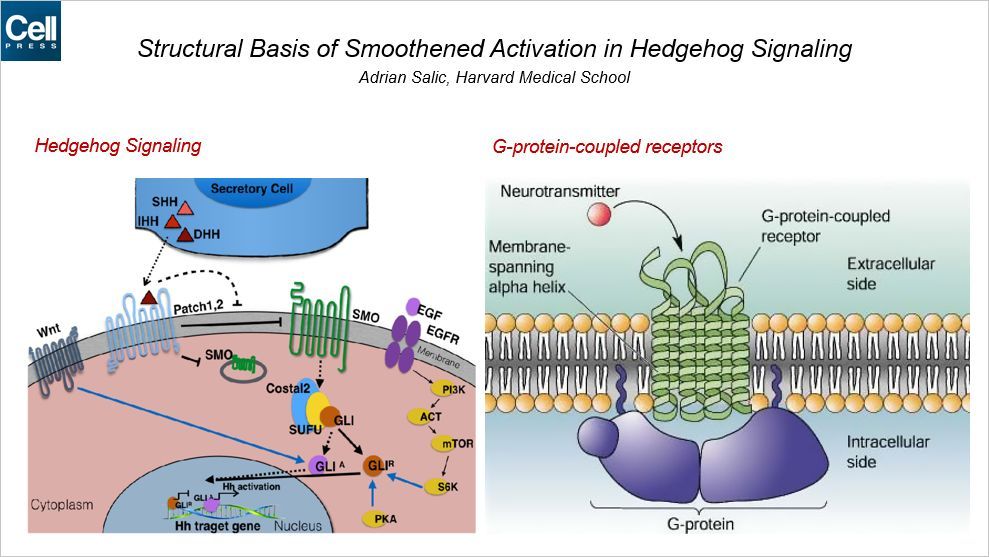
很多动物具有对称结构,比如人、小鼠等。Hedgehog 信号通路对于人的这种对称结构的形成具有重要的作用。最开始人们在果蝇中研究体节形成的关键的基因,科学家们发现当敲除一种基因后,果蝇的胚胎会发育成小的刺状的小球,就像刺猬一样,因此这种蛋白命名为刺猬,这条通路命名为刺猬信号通路。
Hedgehog 蛋白家族主要包含五个成员,它们可以与细胞膜蛋白结合发挥作用。这其中关键的两个膜蛋白分别为 Smoothened(SMO)和 PATCHED(PTCH)。其中 SMO 为 G 蛋白偶联受体。GPCR 最重要的一个特征就是具有七个跨膜的结构域。
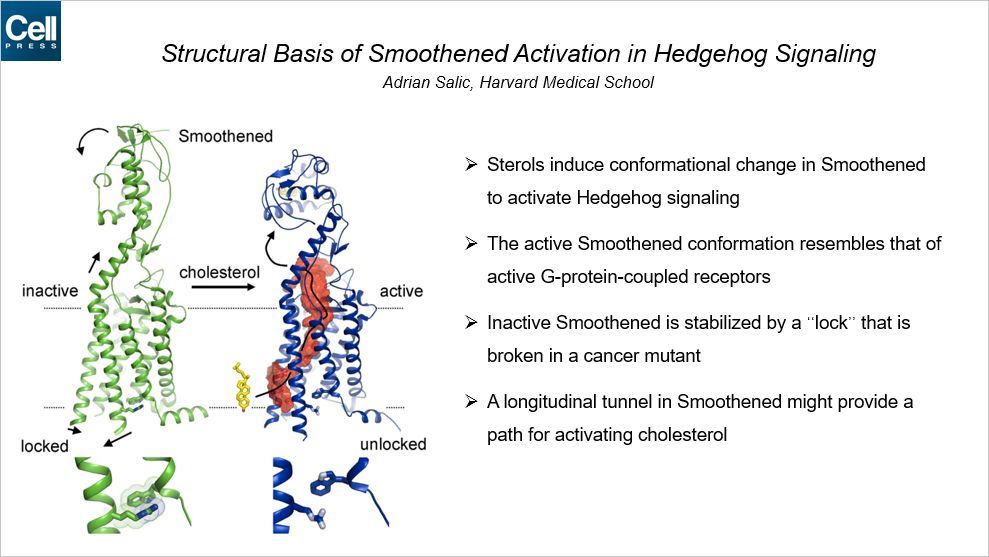
胆固醇结合到 SMO 位于细胞外的富含半胱氨酸的结构域进而激活 SMO。这种相互作用如何影响跨膜结构域的变化以及 SMO 激活?
本文报道了固醇激活的 SMO 的晶体结构。CRD 结构域经历了剧烈的方向调换,通过改变构象导致跨膜结构域采用一种类似于激活的 G-蛋白偶联受体的构象。SMO 含有一个独特的抑制性的 π-阳离子锁,一旦 SMO 被激活,这种锁就被破坏掉,而且在活性致癌突变体中也被破坏。SMO 激活打开了一个疏水的通道,提示胆固醇从内膜移动到 CRD。所有的 SMO 拮抗剂结合到跨膜结构域,封锁通道的打开,但是环巴胺也可以结合到 CRD,诱导活性的跨膜构象。这些结果定义了一种新的 SMO 激活和抑制的机制。
主题六:Pathogen and host interaction
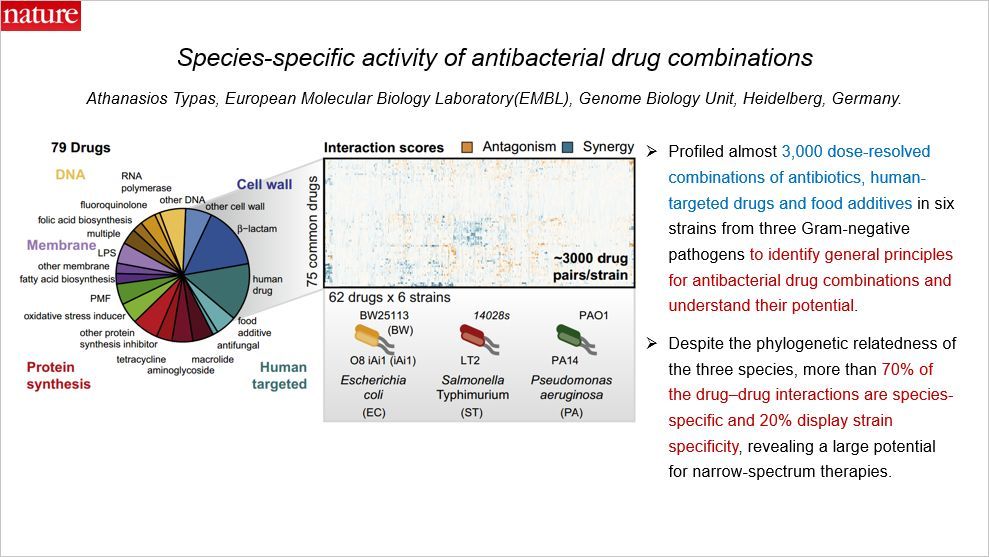
这篇文章是关于抗生素药物的组合使用策略的,通讯作者来自德国。抗生素耐药的蔓延已经成为一个严重的公共健康问题,这让很多此前一度被战胜的疾病又变得致命。药物组合可以战胜多种药物抗性的细菌感染,但是其背后的机制还并不清楚,而且在临床上还没有广泛推广。
这篇文章在三类革兰氏阴性细菌(大肠杆菌、鼠伤寒沙门菌、绿脓假单胞菌)的 6 个种系中测试了 3000 种抗生素、人类靶向药物和食品添加剂(比如香草醛,其也具有一定的抗菌作用)的组合,希望鉴定抗菌药物组合的基本原理,理解这些组合的抗菌潜力。
尽管三个物种存在系统发育相关性,但是在作者所检测的药物组合中,超过 70% 具有物种特异性,20% 显示出种系特异性,提示这些窄谱治疗组合具有巨大的应用潜力。总的来看,药物间拮抗作用比协同增效效应更加常见,而且这种拮抗作用几乎全部出现在靶向不同细胞过程的药物之间;相反,协同增效效应更加保守而且主要出现在靶向同一过程的药物中。此外,这篇文章的结果还显示一些药物组合的协同增效作用在对抗病菌感染过程中显示出较强的效果。
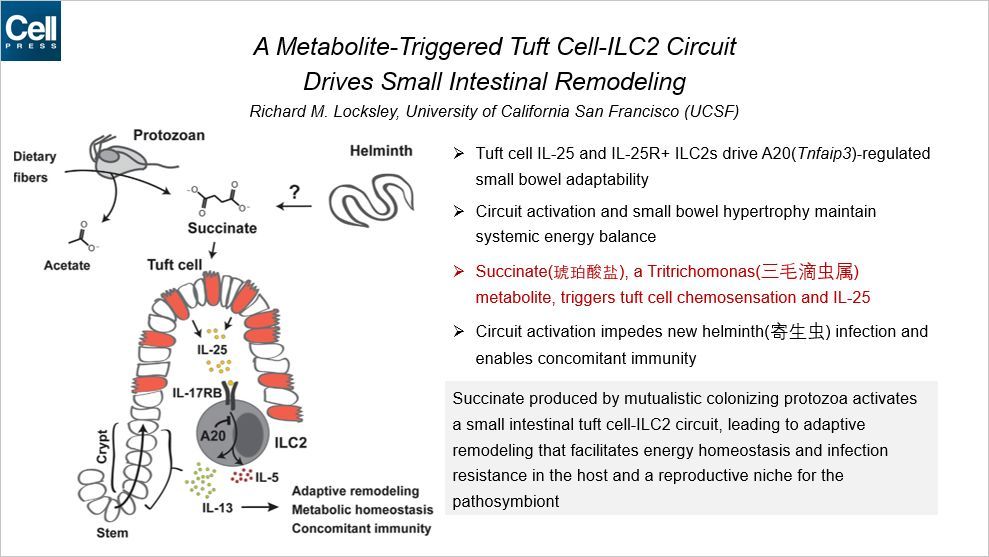
本文聚焦的是病原体的代谢物对小肠簇细胞和小肠的影响,通讯作者来自 UCSF。簇细胞(Tuft Cell)是小肠内的一类能感受化学信号的细胞。炎症性 2 型天然淋巴细胞(group 2 innate lymphoid cell,ILC2)可分泌二型细胞因子响应寄生虫的感染。
簇细胞-ILC2 环路,通过簇细胞化学感应样(chemosensory-like)的感知和 IL-25 介导的固有层 ILC2 的激活,介导肠上皮对肠道寄生虫和原生生物的响应。小肠 ICL2 组成性地表达 IL-25 受体,而IL-25 受体受到 A20(Tnfaip3)的负向调控。ICL2 中 A20 的缺乏同时触发环路和促进适应性的小肠变长和发生重构。而毛滴虫的代谢物琥珀酸盐可以触发簇细胞化学响应和 IL-25 分泌。环路的激活阻碍了新的寄生虫感染,产生了伴随免疫。
主题七:Methodology
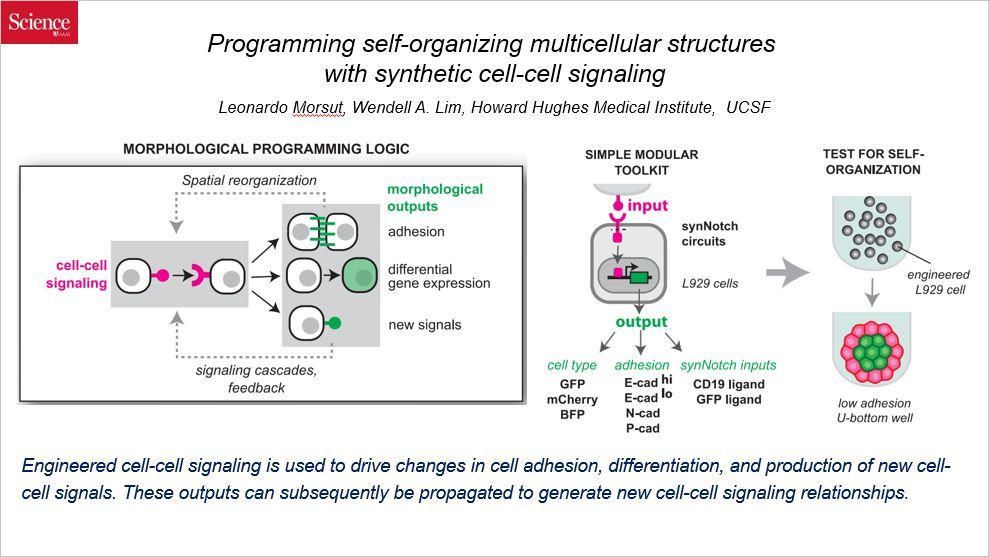
人类有很多器官,比如肝脏、肾脏,肝脏细胞和肾脏细胞如何组装成具有特定形状和功能的器官的,这是长期以来极具挑战性的课题。不难理解的是,细胞与细胞间的交流对于特定组织的形成是至关重要的。
这篇文章通过合成生物学的方法,人为地构造了名为“synNotch 合成的 notch 受体系统(syntheticnotch receptor system)”的细胞间的信号网络,其中特定的细胞-细胞间连接诱导了细胞黏附中钙粘着蛋白的变化。尽管这套系统还很简单,这些小的细胞间的程序足以产生细胞聚集体,而且还具有自然发育系统的一些标志,比如很强的自组织成多结构域结构的能力、精心编排的连续组装、细胞类型多样化、对称破裂(symmetry breaking)以及受伤后的再生能力。这套人工网络的驱动复杂结构形成的能力,证实了构建细胞间信号连接的能力:信号诱导的空间重新组织改变了每个细胞所接受的局部信号,造成了细胞命运分支的迭代循环。这些结果提供了多细胞进化的新观点,揭示了工程化定制的自组织的组织或材料的潜力。
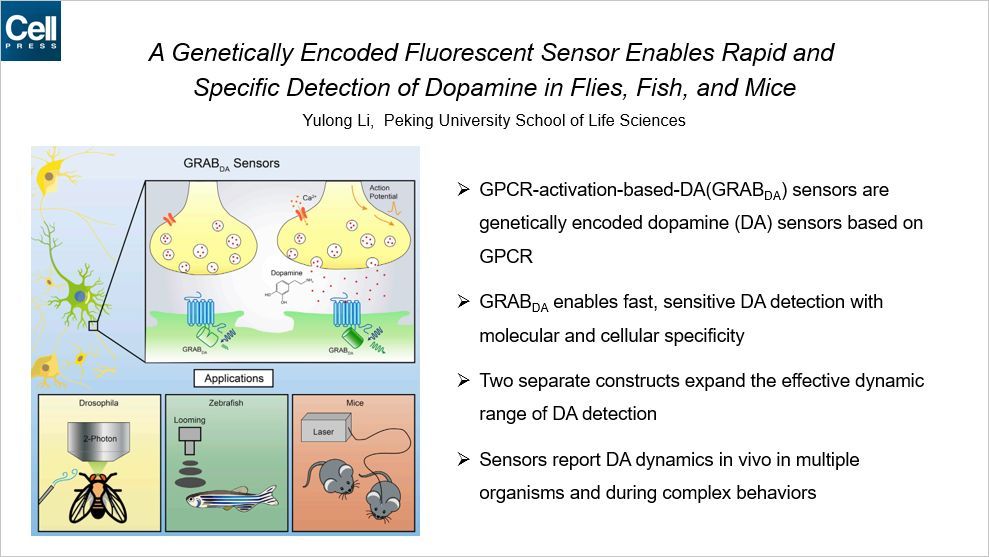
这篇文章来自与北大生科院的李毓龙教授。多巴胺是一种单胺类神经递质,是神经信号传递的分子基础。这篇文章中李毓龙课题组开发了一种基于 G-蛋白耦联受体的,基因编码的多巴胺感知器。利用这套系统,可以快速高灵敏度地检测多巴胺的水平。
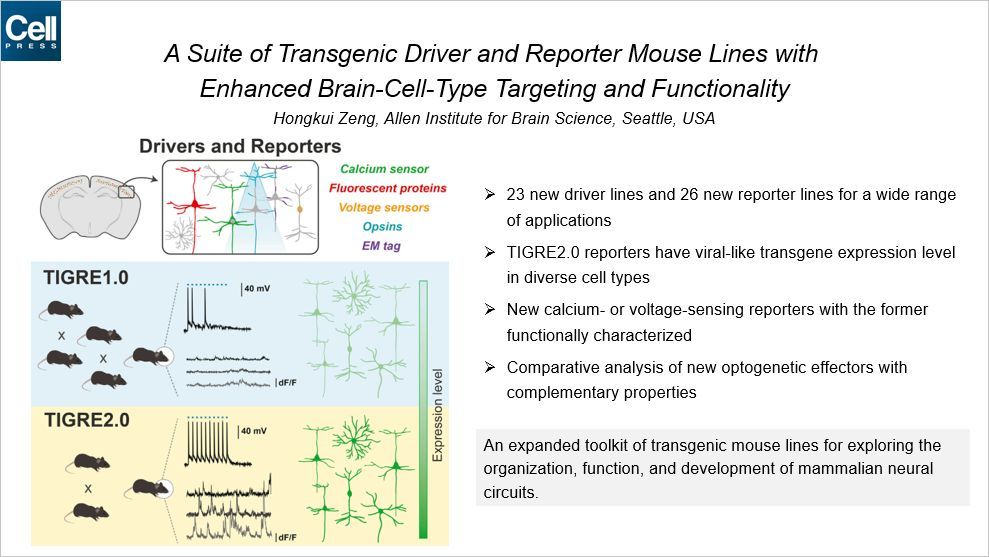
这套增强版的转基因小鼠工具盒 TIGRE2.0 包含了 23 个新的驱动谱系和 26 个新的受体谱系,可以感知钙离子、荧光蛋白、电压等。TIGRE2.0 相比于此前的版本,灵敏度更高。
说在最后:其实从上面的解读来看,我不得不承认,有部分文章还是没有看明白。主要是缺少背景知识,如果想要啃透,需要花费更多的时间,显然这是不理智的。这里提到的通过提问来快速抓住文章要点的方法,希望对大家有用。当然,前辈或同行有更多好的方法,欢迎留言分享。





